溶酶體的生物發生[細胞生物學]
發布時間:2011-10-18 共1頁
溶酶體的生物發生
溶酶體的形成是一個相當復雜的過程, 涉及的細胞器有內質網、高爾基體和內體等。比較清楚的是甘露糖-6-磷酸途徑(mannose 6-phosphate sorting pathway):溶酶體的酶類在內質網上起始合成, 跨膜進入內質網的腔, 在順面高爾基體帶上甘露糖6-磷酸標記后在高爾基體反面網絡形成溶酶體分泌小泡, 最后還要通過脫磷酸才成為成熟的溶酶體(圖9-44)。
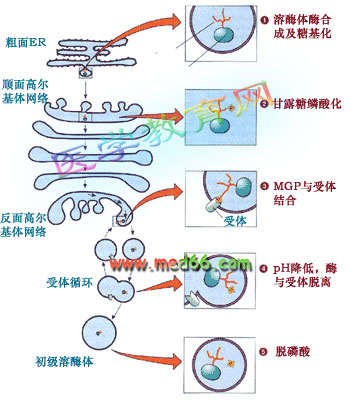
圖9-44 溶酶體的酶尋靶過程、涉及的細胞器及機理
大多數溶酶體的酶在寡糖鏈上含有甘露糖, 在順面高爾基網絡轉變成甘露糖-6-磷酸。新形成的溶酶體的酶通過高爾基復合體,在高爾基體反面網絡與膜受體結合后被包進溶酶體分泌小泡,通過出芽形成自由的分泌泡。通過H+-質子泵調節溶酶體分泌小泡中的pH,使溶酶體的酶同受體脫離,受體再循環, 溶酶體酶脫磷酸后成為成熟的初級溶酶體。
■ 溶酶體酶蛋白的M6P標記
研究發現,溶酶體的酶上都有一個特殊的標記∶6-磷酸甘露糖(mannose 6-phosphate, M6P)。這一標記是溶酶體酶合成后在粗面內質網和高爾基體通過糖基化和磷酸化添加上去的。
● 糖基化
溶酶體酶蛋白在膜旁核糖體上合成,進入內質網后進行N-連接糖基化, 經加工后形成帶有8個甘露糖殘基和2個N-乙酰葡萄糖胺殘基的糖蛋白轉運到高爾基體。
● 信號斑(signal patch)
信號斑是溶酶體酶蛋白多肽形成的一個特殊的三維結構, 它是由三段信號序列構成的, 可被磷酸轉移酶特異性識別(圖9-45)。
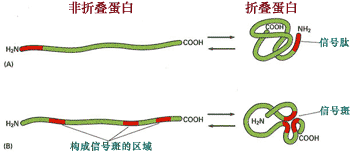
圖 9-45 信號斑
溶酶體蛋白的多肽上有三段信號序列, 通過折疊,三個信號序列相互靠近形成信號斑結構。
● 甘露糖磷酸化的酶
將磷酸基團添加到溶酶體酶的甘露糖的第六位碳上的反應是由兩種酶催化的,一種酶是N-乙酰葡萄糖胺磷酸轉移酶(N-acetyglucosamine phosphotransferase),另一個酶是N-乙酰葡萄糖苷酶, 功能是釋放N-乙酰葡萄糖胺(46)。
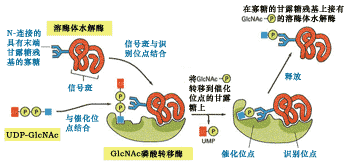
圖9-46 溶酶體酶蛋白信號斑與磷酸化酶相互作用
順面高爾基體中的N-乙酰葡萄糖胺磷酸轉移酶有兩個功能位點,一個識別位點能夠識別信號斑并與之結合;催化位點與高甘露糖N-連接的寡聚糖以及UDP-GlcNAc結合。
● 磷酸化反應
反應中磷酸基的供體是UDP N-乙酰葡萄糖胺(N-acetyglucosamine,GlcNAc), 甘露糖殘基磷酸化的位點是第六位碳原子(圖9-47)。每個溶酶體酶蛋白至少有一個甘露糖殘基被磷酸化。
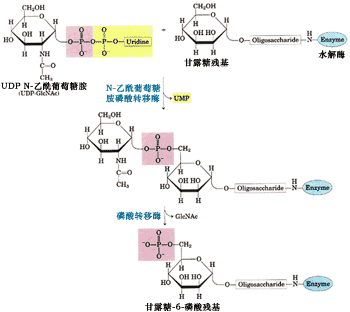
圖9-47 溶酶體酶蛋白甘露糖殘基磷酸化的生化反應
■ 溶酶體酶的M6P分選途徑
這一途徑的兩個關鍵是:M6P標記和M6P受體蛋白。
● M6P受體蛋白(M6P receptor protein)
M6P受體蛋白是反面高爾基網絡上的膜整合蛋白, 能夠識別溶酶體水解酶上的M6P信號并與之結合, 從而將溶酶體的酶蛋白分選出來。
M6P受體蛋白主要存在于高爾基體的反面網絡,但在一些動物細胞的質膜中發現有很多M6P受體蛋白的存在, 這是細胞的一種保護機制, 可防止溶酶體的酶不正確地分泌到細胞外。
● 分選過程
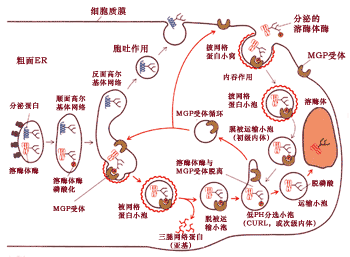
圖9-48 綜合了溶酶體酶的甘露糖6-磷酸分選途徑和溶酶體形成的主要過程。
溶酶體酶前體從粗面內質網轉移到順面高爾基體, 并進行甘露糖殘基的磷酸化。在反面高爾基網絡,磷酸化的酶同M6P受體結合,通過該受體將溶酶體的酶包裝到由纖維狀網格蛋白包被的小泡中, 然后網格蛋白外被很快解體。無包被的運輸小泡很快與次級內體融合, 由于次級內體中pH呈酸性, 致使磷酸化的酶與M6P受體脫離, 接著脫磷酸。通過次級內體的分選作用。 溶酶體的酶進入從次級內體出芽形成的運輸小泡, 接著同溶酶體融合完成溶酶體酶的傳遞過程。而受體重新回到高爾基體再利用。為簡化起見, 圖中只標出酶蛋白的一個糖基化位點, 實際上大多數糖蛋白具有多個糖基化的位點。M6P受體也位于細胞質膜中, 它能夠同偶爾分泌到細胞外磷酸化的溶酶體酶結合, 并形成由網格蛋白包被的運輸泡, 最后同樣被傳遞給溶酶體。
●影響M6P分選的因素
Brown 和Farquhar 發現用胺離子(NH4+)處理細胞能夠干擾溶酶體的分選機制。當溶酶體中胺離子濃度升高時會使溶酶體中的pH升高, 這樣, 溶酶體的酶就不能同M6P受體脫離, 從而影響了M6P受體回到高爾基體再循環。其結果,由于高爾基體反面網絡中M6P受體的不足, 溶酶體的酶就會分泌到細胞外而不是被包裝到溶酶體分泌小泡。如果解除NH4+的作用, 使M6P受體得以釋放和再循環, 溶酶體的分選恢復正常。
●M6P分選途徑的特點:
①M6P作為分選信號; ②包埋在高爾基體中的受體能夠被網格蛋白包裝成分泌小泡; ③出芽形成的溶酶體酶的運輸小泡只同酸性的次級內體融合; ④通過次級內體的分選作用使受體再循環。
■ M6P分選途徑的發現
M6P分選途徑是通過對一類遺傳病: 稱為溶酶體貯積癥(lysosomal storage diseases)的研究發現的,此類遺傳病是由于溶酶體中缺少一種或幾種酶所致。
■ 內體(endosome)
內體有初級內體(early endosome)和次級內體(late endosome)之分, 內體的主要特征是酸性的、不含溶酶體酶的小囊泡。初級內體是由于細胞的內吞作用而形成的含有內吞物質的膜結合的細胞器。
次級內體中的pH呈酸性, 且具有分揀作用。內體膜上具有ATPase-H+ 質子泵,利用H+ 質子的濃度,保證了內部pH的酸性(圖9-49)。
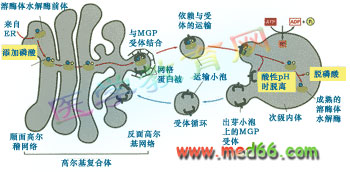
圖9-49 溶酶體酶運輸小泡與次級內體的融合及次級內體的分選作用
■ 溶酶體形成的非M6P途徑
M6P途徑是溶酶體酶分選的主要途徑,但不是惟一的途徑, 這主要是通過對一種遺傳病的研究發現的。
● 粘脂病(mucolipidosis)是一種遺傳病,這種病人不能使甘露糖磷酸化。病人的成纖維細胞中含有很多細胞質小泡,小泡中有大量的未被消化的大分子, 這些大分子在正常情況下是由溶酶體降解的。由于甘露糖不被磷酸化,意味著大多數溶酶體酶不能正確地進入溶酶體。
● 非磷酸化的溶酶體酶
在粘脂病的病人細胞的溶酶體中發現有未被磷酸化的水解酶, 推測這些酶是通過非M6P依賴性的分選途徑進入溶酶體的。這種推測從I-細胞病人的肝細胞溶酶體酶分析中得到證實。I-細胞病人的肝細胞中溶酶體的酶也沒有M6P標記,但是能夠進入溶酶體, 這就說明這些溶酶體必然是通過非M6P依賴的途徑進入溶酶體的,但機理尚不清楚。
● 非M6P途徑的可能方式
兩種可能:一是作為膜蛋白,合成時就插在膜上;另一種可能就是作為前體合成并結合在膜上, 進入溶酶體膜后水解釋放到溶酶體腔中。
請舉例說明溶酶體酶進入溶酶體的非M6P途徑的可能方式。
■ 溶酶體與疾病
已知有不少疾病與溶酶體有關, 如貯積癥(storage diseases)、I-細胞病、矽肺、類風濕性關節炎等。
● 矽肺病(silicosis)
● Ⅱ型糖原貯積癥(glycogen storage disease type Ⅱ)
● 休克(shock)
